


Mikroelektronik indlejret i levende vandmænd forbedrer fremdriften
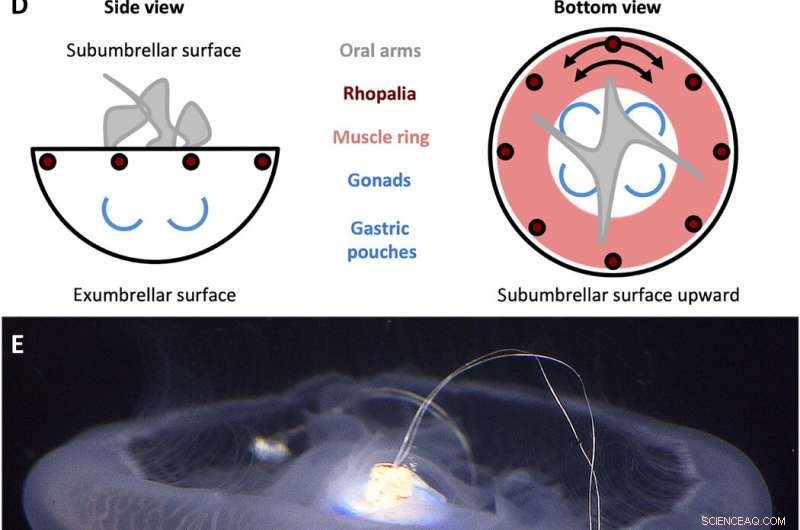
A. aurita svømmecontroller design. (A) Firkantbølgesignal genereret af svømmecontrolleren med en amplitude (A) på 3,7 V og en pulsbredde (T) på 10 ms, indstillet til frekvenser (f) på 0,25, 0,38, 0,50, 0,62, 0,75, 0,88, og 1,00 Hz. (B) Svømmecontrollerkomponenter. Huset inkluderer (i) en polypropylenhætte med en træstift, der indlejres i klokkecentret, og (ii) en plastikfilm til at vandtætte huset, begge forskudt med rustfrit stål og korkvægte for at holde enheden omtrent neutralt flydende. Mikroelektronik omfatter (iii) en TinyLily mini-processor, (iv) lithium polymer batteri, og (v) to platin-spidselektroder med LED'er til visuelt at indikere stimulering. (C) Fuldt samlet enhed, med processor og batteri indkapslet i huset. (D) Forenklede skemaer af A. auritas anatomi, fremhæve subumbrellar (øverst) og exumbrellar (nederste) overflader, rhopalia, muskel ring, og perifer muskelfiberorientering, mundtlige arme, og gonader/maveposer. (E) Svømmecontroller (inaktiv) indlejret i en fritsvømmende vandmand, klokkeorienteret subparaply side opad, med træstiften indsat i manubrium og to elektroder indlejret i muskel- og mesoglealt væv nær klokkekanten. Billedkreditter for (B), (C), og (E):Nicole W. Xu, Stanford University. Kredit:Science Advances, doi:10.1126/sciadv.aaz3194
Forskere i robotmaterialer sigter mod kunstigt at kontrollere dyrs bevægelse for at løse de eksisterende udfordringer med aktivering, kontrol- og effektkrav i blød robotteknologi. I en ny rapport i Videnskabens fremskridt , Nicole W. Xu og John O. Dabiri ved afdelingerne for bioingeniør, civil- og miljøteknik og maskinteknik ved Stanford University præsenterede en biohybrid robot, der brugte mikroelektronik ombord til at fremkalde svømning i levende vandmænd. De målte evnen til at forbedre fremdriften væsentligt ved at drive kropssammentrækninger ved et optimalt frekvensområde hurtigere end naturlig adfærd. Manøvren øgede svømmehastigheden næsten tre gange, dog med kun en dobbelt stigning i dyrets metaboliske forbrug og 10 mW ekstern effekttilførsel til mikroelektronikken. Biohybridrobotten brugte 10 til 1000 gange mindre ekstern strøm pr. masse end tidligere rapporterede vandrobotter. Muligheden kan forbedre ydeevneomfanget af biohybride robotter i forhold til native ydeevne, med potentielle anvendelser som biohybride havovervågningsrobotter.
Vandmænd er en overbevisende modelorganisme til at danne energieffektive undervandsfartøjer på grund af deres lave transportomkostninger (COT). Eksisterende biomimetiske robotter af svømmende dyr, der udelukkende er bygget af konstruerede materialer, kan opnå hastigheder, der kan sammenlignes med naturlige dyr, men med størrelsesordener mindre effektive end vandmænd. Biohybride vandmandsrobotter kan derfor integrere levende dyr for at løse eksisterende udfordringer med blød robotteknologi. Forskere kan bruge vandmandsstrukturen til aktivering og løse strømkrav ved at udforske naturlig fodringsadfærd, hvor de udvinder kemisk energi fra byttedyr in situ. Tilgangen kan også tillade genopretning fra skade via naturlige sårhelingsprocesser, der er iboende for dyret, kontrollere dyrs bevægelse og tillade yderligere undersøgelser af levende organismers biomekanik i brugerkontrollerede eksperimenter. I dette studie, Xu og Dabiri brugte et system af mikroelektronik til eksternt at kontrollere en levende vandmand og danne en biohybrid robot til at fremme videnskab og teknik inden for akvatisk bevægelse.
For at aktivere vandmænd som et naturligt stillads, holdet brugte dyrets eget basale stofskifte til at reducere yderligere kraftbehov og udnyttede dets muskler til aktivering, mens de stolede på selvhelbredende og vævsregenererende egenskaber for øget skadetolerance. Holdet antog, at øgede klokkekontraktionsfrekvenser for vandmænd kunne øge svømmehastigheden op til en grænse. De styrede derfor eksternt frekvensen af pulser hos fritsvømmende dyr ved at måle svømmehastigheden og iltindtaget for at beregne transportomkostningerne (COT) og teste deres arbejdshypotese. Tidligere var sådanne undersøgelser kun mulige gennem beregningsmæssige eller teoretiske modeller.
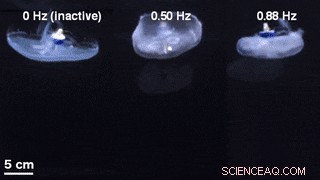
En sammenligning af klokkegeometrier for ustimuleret svømning med en inaktiv svømmecontroller indlejret (venstre) og eksternt styret svømning ved 0,50 Hz (midten) og 0,88 Hz (højre). Kredit:Science Advances, doi:10.1126/sciadv.aaz3194
Xu et al. udvalgte Aurelia aurita som en modelorganisme; en oblate art af vandmænd, der indeholder en fleksibel mesogleal klokke og et enkelt lag af koronale og radiale muskler, der beklæder subumbrellaoverfladen. For at svømme, organismerne trak musklerne sammen for at mindske rumfanget i det subumbrella hulrum og udstøde vand for at give en drivkraft sideløbende med yderligere bidrag fra passiv energigenvinding og sugebaseret fremdrift. For at starte disse muskelsammentrækninger, vandmændene aktiverede enhver af dens lette pacemakere placeret i sensororganerne kendt som rhopalia langs klokkekanten. Disse nerveklynger aktiverede hele det motoriske nervenett for at forårsage tovejs muskelbølgeudbredelse, der stammede fra de aktiverede pacemakere under naturlig udbredelse.
Robotdesignintegration i levende vandmænd og enhedsvalidering
Forskerne konstruerede først en bærbar, selvstændig mikroelektronisk svømmecontroller til at generere en firkantet pulsbølge og stimulere muskelsammentrækninger fra 0,25 Hz til 1,00 Hz. De sammensatte controlleren med en TinyLily mini-processor og en 10 mAh lithium polymer celle. For visuelt at bekræfte det elektriske signal, Xu et al. forbundet ledningerne i serie til TinyLily lysemitterende dioder (LED'er). Derefter indsatte de elektroder bilateralt i subumbrellavævet og holdt systemet naturligt flydende med rustfri stålskiver og kork. For at validere, at svømmecontrolleren eksternt kunne kontrollere vandmænds klokkesammentrækninger, forskerne udviklede en metode til at spore bevægelsen af klokkekanten. For det, de gennemførte tre sæt eksperimenter, (1) at observere endogene sammentrækninger af organismen i fravær af forstyrrelser, (2) at observere, om mekanisk indlejring af inaktive elektroder påvirkede naturlig dyreadfærd og (3) at teste stimuleringsprotokoller for at bekræfte eksternt drevne sammentrækninger.
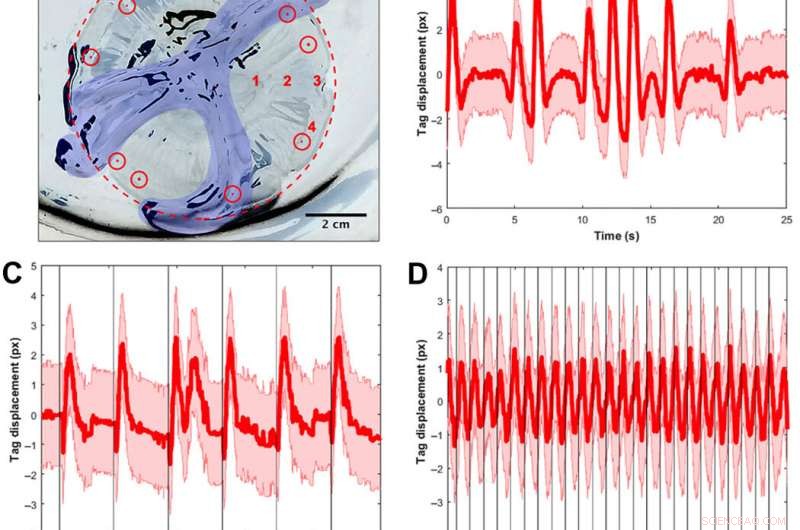
Signalvalidering ved hjælp af visuelle tags og frekvensspektre til at spore muskelsammentrækninger. (A) A. aurita medusae (n =10, 8,0 til 10,0 cm i diameter) blev anbragt under paraplyoverfladen op i en plade uden havvand til forsøg med begrænset muskelstimulering (elektrode ikke vist). Billedet er vendt om, så klokken og pladen er hvide, og sorte områder er refleksioner af lys fra dyrevæv og pladen. For klarhedens skyld, klokkens margin er skitseret i en rød stiplet cirkel, og mundarmene er farvet i blåt. Synlige implantat-elastomer-tags (vist som farvede røde prikker i røde cirkler) blev injiceret rundt om marginen, og et mærke blev sporet pr. video for at beregne vævsforskydningen som et surrogat for muskelsammentrækninger. Rumlige tests for at bestemme, om elektrodeplacering påvirkede spektrene, blev udført på fire steder, mærket med røde tal:(1) ved siden af maveposerne, (2) midtvejs mellem maveposerne og kanten, (3) ved rhopalia, og (4) i kanten væk fra rhopalia. Alle andre test blev udført på lokation 2. (B) Eksempel tag-forskydning som en funktion af tiden for et dyr uden nogen ekstern stimulus. Den røde linje angiver tyngdepunktsforskydningen, med fejlen beregnet ud fra at antage en halv-pixel usikkerhed ved at finde tyngdepunktet for tagget i hvert billede, over 25 sek. Bemærk den tidsmæssige variation af muskelsammentrækninger, inklusive perioder med regelmæssige pulser og på hinanden følgende hurtige pulser. (C) Eksempel tag forskydning for et dyr med en ekstern stimulus på 0,25 Hz, med hver stimulus visualiseret som en lodret sort linje. Selvom sammentrækninger regelmæssigt følger ydre stimuli, naturlige dyreimpulser forekommer også ved lave frekvenser. Bemærk, for eksempel, dobbeltpulsen efter én stimulus (t ≈ 12 s). (D) Eksempel tag forskydning for et dyr med en ekstern stimulus på 1,00 Hz, med hver stimulus visualiseret som en lodret sort linje. Det samme tidsvindue (25 s) er vist for en rimelig sammenligning med de to foregående plots. Sammentrækninger følger regelmæssigt ydre stimuli. (E) enkeltsidet amplitudespektrum (SSAS) beregnet i gennemsnit for vandmænd uden nogen ekstern stimulus (n =12 for 10 dyr, dvs. 2 vandmænd havde to replikatklip hver). Den røde linje angiver gennemsnittet af normaliseret SSAS for hver replikat, med SD i pink. Toppen af den gennemsnitlige SSAS er på 0,16 Hz. Den fulde bredde ved halv maksimum (FWHM) er 0,24 Hz. (F) Vandmænds respons på en inaktiv elektrode indlejret (n =14 for 10 dyr, dvs. 4 vandmænd havde to replikatklip hver). Toppen af den gennemsnitlige SSAS er på 0,18 Hz. FWHM er 0,16 Hz. Ved at bruge en to-prøve t-test af spidsfrekvenserne for begge grupper, forskellen mellem de to prøver var statistisk insignifikant (P =0,68). (G) Prøve SSAS for en elektrisk stimulus ved 1,00 Hz (n =10 vandmænd for et indgangssignal på 4,2 V og 4,0 ms). Spidsfrekvensen forekommer ved 1,02 Hz, inden for vinduet 0,02, der bruges til at beregne SSAS. Bemærk, at spektret har en skarpere top ved frekvensen af interesse (FWHM på 0,04 Hz), i modsætning til en bredere FWHM i (B) og (C), tilfældene uden nogen ydre stimulus. (H) Konturkort over frekvensresponsen af muskelsammentrækninger til eksterne elektriske stimuli. Hver lodret linje af data (centreret på hvide linjer ved 0,25, 0,50, 0,75, 1.00, 1,20, 1,50, og 2,00 Hz) repræsenterer PSD'en ved én elektrisk indgangsfrekvens, med antallet af testede vandmænd vist ovenfor. Farverne svarer til PSD'ens amplitude, hvor højere værdier vises i gult og lavere værdier i blåt. Den ubrudte røde linje repræsenterer et en-til-en input-output svar, og den stiplede røde linje repræsenterer den rapporterede fysiologiske grænse i henhold til den minimale absolutte refraktære periode for A. aurita muskel (32). Responsive forsøg er defineret ved, om spidsfrekvenserne i PSD'en ligger inden for et vindue på 0,06 Hz af den solide røde kurve. (I) Konturkort over forsøgene, der ikke reagerer. Højere frekvenser op til 90,00 Hz blev også testet med lignende ikke-responsive PSD'er. Billedkredit for (A):Nicole W. Xu, Stanford University. Kredit:Science Advances, doi:10.1126/sciadv.aaz3194
De fandt, at naturlig dyreadfærd (eller endogen kontraktion) var uregelmæssig med høj pulsfrekvensvariabilitet - inklusive en gennemsnitlig spidsfrekvens på 0,16 Hz. En inaktiv elektrode ændrede ikke signifikant frekvensspektrene, mens eksternt drevne kontraktioner viste en fysiologisk grænse for vandmænds muskelkontraktioner mellem 1,4 Hz til 1,5 Hz. Holdet gennemførte svømmeforsøg med det implanterede system i en saltvandstank og normaliserede de målte svømmehastigheder for at tage højde for variation i dyrestørrelse. De skalerede den normaliserede svømmehastighed med gennemsnittet af den normaliserede hastighed i fravær af stimulering (dvs. 0 Hz) for at bestemme forstærkningsfaktoren. Den maksimale forbedringsfaktor var op til 2,8 gange dyrenes naturlige svømmehastighed, dvs. svømmehastigheden blev forbedret op til 2,8 gange ved hjælp af indbygget mikroelektronik.
Meget effektivt enheds strømforbrug
De kunstigt kontrollerede vandmænd krævede ekstern strøm fra det mikroelektroniske system og indre kraft fra dyrenes eget stofskifte. Når der køres med stigende frekvenser, det mikroelektroniske system af den biohybride robotvandmand forbrugte større watt pr. kg. Imidlertid, sammenlignet med eksisterende robotter, denne biohybrid robot brugte op til 1000 gange mindre ekstern strøm. Xu et al. sammenlignede denne prototype med medusoid- og robotstrålen fremstillet af rotte-kardiomyocytter udsået på siliciumstilladser, og med rent mekaniske robotter samt autonome undervandsfartøjer (AUV'er). Ud over de omkostningseffektive fordele ved lavt eksternt strømforbrug pr. masse af biohybridrobotten, det mikroelektroniske system koster kun mindre end $20 fra kommercielt tilgængelige komponenter. Elektrolokationen var også uspecifik, og dyrene kom sig straks efter forsøgene.
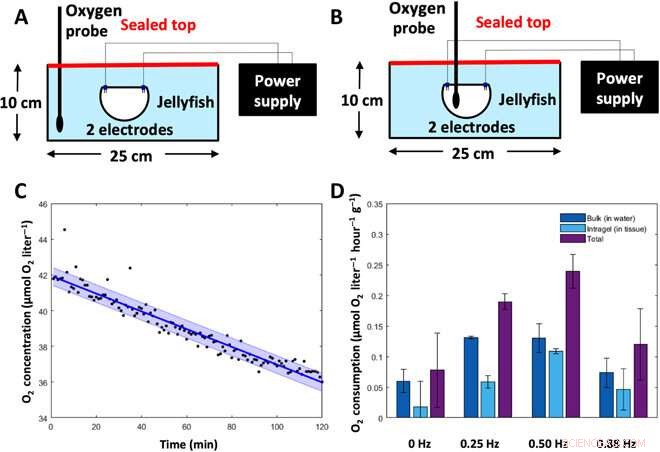
Metaboliske hastighedsforsøg. For at bestemme vandmændenes stofskiftehastighed, oxygen concentrations were measured in animal tissue and the surrounding water and then converted into energy expenditure. (A) Experimental setup to measure bulk dissolved oxygen concentrations (in the water). Animals were placed subumbrellar surface upward in a sealed glass dish filled with 2 liters of artificial seawater, with two electrodes for frequency-driven cases. Oxygen levels in the water were measured using a MicroOptode oxygen probe. (B) Experimental setup to measure intragel oxygen concentrations (in the tissue). Animals were placed subumbrellar surface upward in a sealed glass dish filled with 2 liters of artificial seawater, with two electrodes for frequency-driven cases. Intragel oxygen levels were measured using a MicroOptode oxygen probe embedded into the tissue. (C) Representative plot of oxygen concentrations over time, measured from the MicroOptode. This example shows measurements of bulk oxygen levels in the water surrounding an animal with a swim controller–driven frequency of 1.00 Hz. Individual data points are shown in black, the best-fit line is shown in dark blue, and the SD is shown in the light blue shaded region. (D) Oxygen consumption rates of the surrounding water (dark blue), within animal tissue (light blue), and total (sum of the water and tissue measurements, purple) were calculated over a 6- to 8-hour period (n =7 animals).Credit:Science Advances, doi:10.1126/sciadv.aaz3194
The new capability of external control allowed Xu et al. to address the relationship between swimming frequency and metabolic rate. Oxygen consumption rates followed a similar pattern to enhanced swimming speeds, and the scientists calculated the equivalent cost of transport using both experimental metabolic rates and experimental swimming speeds. The COT increased at mid-range frequencies and decreased at high external stimulation frequencies. The results showed that enhanced jellyfish swimming did not cause undue cost to the metabolism or health of the animal.
The main robotic limit of the study was the power requirement of the microelectronic system relative to animal versus microelectronic power needs. Further improvement to microelectronics can decrease the energetic costs and extended studies can also strive to minimize endogenous animal contractions without harming the organism to improve controllability of live-animal-based biohybrid robots. The artificial control of jellyfish can expand ocean monitoring techniques with improved controllability by incorporating microelectronic sensors to leverage the existing tagging technology.
© 2020 Science X Network
 Varme artikler
Varme artikler
-
 KLM erstatter Bruxelles-flyvning med tog for at reducere emissionerneKLM vil fra marts 2020 reducere antallet af flyvninger fra Amsterdam til Bruxelles fra fem til fire i et forsøg på at reducere sin miljøbelastning Er det en fugl, er det et fly? Ingen, siger det h
KLM erstatter Bruxelles-flyvning med tog for at reducere emissionerneKLM vil fra marts 2020 reducere antallet af flyvninger fra Amsterdam til Bruxelles fra fem til fire i et forsøg på at reducere sin miljøbelastning Er det en fugl, er det et fly? Ingen, siger det h -
 Privat, stabil og landede:Mød Tor Browser 8.5En Tor-browser er ankommet som en stabil udgivelse til Android. Ultraprivat. Og nogle Tor-ønskelister vil sige, endelig. Indtil for nylig, det var kun tilgængeligt på stationære enheder. Nu, den stabi
Privat, stabil og landede:Mød Tor Browser 8.5En Tor-browser er ankommet som en stabil udgivelse til Android. Ultraprivat. Og nogle Tor-ønskelister vil sige, endelig. Indtil for nylig, det var kun tilgængeligt på stationære enheder. Nu, den stabi -
 At gøre flyvning faktisk bæredygtigtDet nye solminiraffinaderi, der ligger på taget af ETHs Machine Laboratory-bygning i Zürich, kan producere klimaneutrale brændstoffer til bæredygtig luftfart i fremtiden. Kredit:Tricklabor / ETH Züric
At gøre flyvning faktisk bæredygtigtDet nye solminiraffinaderi, der ligger på taget af ETHs Machine Laboratory-bygning i Zürich, kan producere klimaneutrale brændstoffer til bæredygtig luftfart i fremtiden. Kredit:Tricklabor / ETH Züric -
 At misbruge en robot skader den ikke, men det kan gøre dig til en grusommere personKredit:CC0 Public Domain Udspillet i et dystopisk 2019, sci-fi-klassikeren Blade Runner udforsker, hvordan kunstige mennesker kan påvirke vores menneskelighed. Harrison Fords karakter oplever stær
At misbruge en robot skader den ikke, men det kan gøre dig til en grusommere personKredit:CC0 Public Domain Udspillet i et dystopisk 2019, sci-fi-klassikeren Blade Runner udforsker, hvordan kunstige mennesker kan påvirke vores menneskelighed. Harrison Fords karakter oplever stær
- Uovertruffen nøjagtighed inden for kvanteelektrodynamik:Kæmpespring mod at løse protonladningsrad…
- Amazonfloden pirater vand fra nabolandet Rio Orinoco
- Efterligner træets ultrastruktur med 3D-print til grønne produkter
- Dråbespredning fra mennesker følger ikke altid luftstrømmen
- Levering af kræft-billeddannende isotoper af høj kvalitet
- Spændingsstigninger på op til 25% observeret i tætpakkede nanotråde