


Forskere udforsker, hvordan genomisk integritet bevares i dobbeltstrengsbrud
For at forhindre sådanne katastrofale hændelser har celler udviklet indviklede mekanismer til at reparere DNA-dobbeltstrengsbrud. Disse mekanismer omfatter to hovedveje:homolog rekombination (HR) og ikke-homolog endesammenføjning (NHEJ).
Homolog rekombination anvender en homolog DNA-sekvens som skabelon til at reparere det ødelagte DNA. Denne vej er meget nøjagtig og forekommer primært under S- og G2-faserne af cellecyklussen, når et søsterkromatid er tilgængeligt som skabelon.
Ikke-homolog endesammenføjning ligerer på den anden side direkte de knækkede DNA-ender uden behov for en skabelon. Selvom den er hurtigere og mindre afhængig af cellecyklusstadiet, er denne vej mere tilbøjelig til fejl og kan resultere i små indsættelser eller sletninger på reparationsstedet.
Valget mellem HR og NHEJ er påvirket af flere faktorer, herunder tilgængeligheden af en homolog skabelon og cellecyklusstadiet. Generelt foretrækkes HR, når en homolog sekvens er til stede, og cellen er i S- eller G2-fasen. I modsætning hertil er NHEJ hyppigere ansat, når der ikke er nogen tilgængelig skabelon eller i hurtigt delende celler, hvor HR er mindre effektiv.
Det er værd at bemærke, at ud over disse to hovedveje kan andre mekanismer bidrage til reparationen af DNA-dobbeltstrengsbrud, herunder alternativ endesammenføjning og enkeltstrengsannealing.
At forstå de mekanismer, der anvendes af celler til at reparere DNA-dobbeltstrengsbrud, er af afgørende betydning på forskellige områder, lige fra kræftforskning til strålebehandling. Ved at målrette mod disse mekanismer kan der udvikles nye terapeutiske tilgange til selektivt at dræbe kræftceller og samtidig skåne sundt væv.
 Varme artikler
Varme artikler
-
 Er John James Audubons tidligere ugerninger kommet hjem for at hvile?Kredit:Det Hvide Hus, Public domain, via Wikimedia Commons Det var en risiko, som sandsynligvis ikke blev opfattet tilbage i 1886, da Forest and Stream-magasinets redaktør George Bird Grinnell, for
Er John James Audubons tidligere ugerninger kommet hjem for at hvile?Kredit:Det Hvide Hus, Public domain, via Wikimedia Commons Det var en risiko, som sandsynligvis ikke blev opfattet tilbage i 1886, da Forest and Stream-magasinets redaktør George Bird Grinnell, for -
 At stole på GPS forhindrer dele af din hjerne i at blive aktiveretBrug trin-for-trin, satellitbaseret navigation kan forhindre dele af din hjerne, der er forbundet med rumlig begrundelse, fra at sparke i højt gear. Klaus Vedfelt/Image Bank/Getty Images Hvornår var
At stole på GPS forhindrer dele af din hjerne i at blive aktiveretBrug trin-for-trin, satellitbaseret navigation kan forhindre dele af din hjerne, der er forbundet med rumlig begrundelse, fra at sparke i højt gear. Klaus Vedfelt/Image Bank/Getty Images Hvornår var -
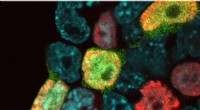 (Gen)erhvervelse af potentialet til at blive altFluorescensbillede af museembryonale stamceller (kerner i blåt) inklusive 2 cellelignende celler (grønne) og den nye population af overgangsceller (rød). Kredit:Helmholtz Zentrum München/IES En ny
(Gen)erhvervelse af potentialet til at blive altFluorescensbillede af museembryonale stamceller (kerner i blåt) inklusive 2 cellelignende celler (grønne) og den nye population af overgangsceller (rød). Kredit:Helmholtz Zentrum München/IES En ny -
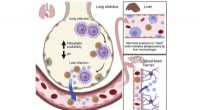 En svamp, der skrumper i størrelse for bedre at inficere hjernenGrafisk abstrakt. Kredit:Cell Host &Microbe (2022). DOI:10.1016/j.chom.2022.08.017 En svamp, der er en almindelig årsag til svampe-meningitis, gennemgår en bemærkelsesværdig transformation, når den
En svamp, der skrumper i størrelse for bedre at inficere hjernenGrafisk abstrakt. Kredit:Cell Host &Microbe (2022). DOI:10.1016/j.chom.2022.08.017 En svamp, der er en almindelig årsag til svampe-meningitis, gennemgår en bemærkelsesværdig transformation, når den
- Hvordan man finder molmassen af KCl
- Kæmpe stjerneudbrud opdaget for første gang
- Australiens bevægelse for at lagre kulstof i jorden er et problem for at tackle klimaændringer
- Sådan reguleres jævnstrøm med modstande
- Hvad er på scenen på CES tech show? Din stemme
- Forskere udvikler måde at forstørre nanostrukturer til lys, fleksible 3D-trykte materialer