


Nøgletræk ved eukaryotisk mRNA:Afdækning og polyadenylering forklaret
Comstock/Stockbyte/Getty Images
DNA lagrer det genetiske plan, der driver cellulære funktioner. I eukaryoter, hvor DNA opholder sig inde i en kerne, skal instruktionerne overføres til cytoplasmaet via messenger RNA (mRNA). Når det først er transskriberet, gennemgår det begyndende mRNA en række enzymatiske redigeringer, der tilføjer væsentlige funktioner, hvilket signalerer, at molekylet er klar til translation.
Capping af mRNA
Den første modifikation, der er fælles for alle eukaryote mRNA'er, er 5'-hætten. Da RNA-polymerase III syntetiserer transkriptet, modificeres 5'-enden efterfølgende af en trio af enzymer, der binder en 7-methylguanylatgruppe. Denne hætte beskytter ikke kun RNA'et mod exonukleaser, men fungerer også som et genkendelsessignal for ribosomer og eksportmaskineri.
Polyadenylering
I den modsatte ende er 3′-terminalen vedhæftet en poly-A-hale af poly(A)-polymerase. Typisk tilføjes 100-250 adenosinrester, en funktion, der forbedrer mRNA-stabilitet og letter dets eksport fra kernen.
Disse ændringers funktionelle betydning
Mens bakterielle mRNA'er mangler både en 5'-hætte og en poly-A-hale, er eukaryote transkripter afhængige af disse strukturer til at regulere nuklear eksport, translationel initiering og RNA-levetid. De tilføjede modifikationer skaber en robust ramme, der sikrer, at kun korrekt behandlede mRNA'er når ribosomet.
Viral manipulation af værts-mRNA
Virus, der inficerer eukaryote celler, kaprer ofte værtens oversættelsesmaskineri. For eksempel koder poliovirus for proteaser, der spalter værts-eIF4G-proteinet, en komponent, der er essentiel for ribosomrekruttering til afgrænsede mRNA'er. Som følge heraf dæmpes cellulære mRNA'er, hvilket tillader virussens uafsluttede RNA at dominere proteinsyntesen - en smart udnyttelse af værtens eget reguleringssystem.
 Varme artikler
Varme artikler
-
 Forskere identificerer gen til at hjælpe hybrid hvede avlKredit:Wikipedia Australske forskere ved University of Adelaide har identificeret et naturligt forekommende hvedegen, der, når den er slukket, eliminerer selvbestøvning, men tillader stadig krydsb
Forskere identificerer gen til at hjælpe hybrid hvede avlKredit:Wikipedia Australske forskere ved University of Adelaide har identificeret et naturligt forekommende hvedegen, der, når den er slukket, eliminerer selvbestøvning, men tillader stadig krydsb -
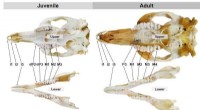 Hvorfor får mennesker to sæt tænder? Disse pungdyr omskriver historien om tandudviklingenBaby- og voksentænder af tammar-wallabyen. Målestok er lig med 1 cm. Kredit:Nasrullah et al. Du får kun 52 tænder i din levetid:20 mælketænder, efterfulgt af 32 voksentænder. Sådan er det ikke for
Hvorfor får mennesker to sæt tænder? Disse pungdyr omskriver historien om tandudviklingenBaby- og voksentænder af tammar-wallabyen. Målestok er lig med 1 cm. Kredit:Nasrullah et al. Du får kun 52 tænder i din levetid:20 mælketænder, efterfulgt af 32 voksentænder. Sådan er det ikke for -
 Neurogenese hos arter af mellemamerikanske fisk afhænger af tilstedeværelsen af prædationsrisik…Braeden Donaldson:Vi ønskede at se, om der var en underliggende neuroplastisk reaktion, der korrelerer med visse adfærdsændringer. Vi fandt ud af, at der var det. Kredit:Concordia University Kemisk
Neurogenese hos arter af mellemamerikanske fisk afhænger af tilstedeværelsen af prædationsrisik…Braeden Donaldson:Vi ønskede at se, om der var en underliggende neuroplastisk reaktion, der korrelerer med visse adfærdsændringer. Vi fandt ud af, at der var det. Kredit:Concordia University Kemisk -
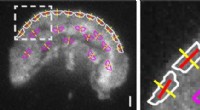 Interne kræfter, der styrer cellemigration, afsløres ved levende cellemikroskopiOrientering af integriner i forkant af en migrerende Jurkat T -celle, analyseret ved hjælp af et fluorescenspolariseret lysmikroskop (øjeblikkeligt FluoPolScope) udviklet på Marine Biological Laborato
Interne kræfter, der styrer cellemigration, afsløres ved levende cellemikroskopiOrientering af integriner i forkant af en migrerende Jurkat T -celle, analyseret ved hjælp af et fluorescenspolariseret lysmikroskop (øjeblikkeligt FluoPolScope) udviklet på Marine Biological Laborato
- Størstedelen af tidlige karriereforskere inden for fysisk videnskab ønsker at offentliggøre åb…
- Har en virus cellemembraner?
- Hvilken type energi er primært forbundet med den tilfældige bevægelse og arrangementspartikler i …
- Svaret på at holde elgbestande sunde? Ulve
- Opretter is og friktion nogen energi?
- Hvordan smelter termisk energioverførsel en isterning ved stuetemperatur?