


Krympende dinosaurer og udviklingen af endotermi hos fugle
Dracoraptor hanigani (teropod) restaureret som et rovdyr, der bor på en kystlinje og ådselæder. Kunstværk af Bob Nicholls (paleocreations.com). Kredit:Naturhistorisk Museum, London. Kredit:Science Advances, doi:10.1126/sciadv.aaw4486
Udviklingen af endotermi (termoregulering ved metaboliske midler) repræsenterer en stor overgang i hvirveldyrs historie. Imidlertid, processen med endotermi-evolution og dens tidslinje hos fugle og pattedyr er fortsat kontroversiel. I en ny rapport vedr Videnskabens fremskridt , Enrico L. Rezende og et team af forskere ved Center of Applied Ecology and Sustainability, og Institute of Environmental and Evolutionary Sciences i Chile, kombineret en varmeoverførselsmodel med kropsstørrelsesdata for theropod. Forskerne rekonstruerede derefter udviklingen af metaboliske hastigheder langs fuglestammen. Resultaterne antydede, at en reduktion i størrelse udgjorde vejen for mindst modstand for endotermi til at udvikle sig - maksimering af termisk nicheudvidelse, samtidig med at omkostningerne ved forhøjede energikrav reduceres.
Som resultat, forskerne antager, at stofskiftet ville være steget med miniaturisering i løbet af tidlig-mellemjuraperioden (ca. 180 til 170 millioner år siden) for at resultere i en gradient af metaboliske niveauer i theropodens fylogeni. Mens basale theropoder kan have udvist lavere stofskiftehastigheder, de nyere ikke-fugle-slægter var sandsynligvis anstændige termoregulatorer med forbedret stofskifte. Analysen gav en foreløbig sekvens i tid af de vigtigste evolutionære overgange, for fremkomsten af små, endotermiske og flyvende fjerdinosaurer.
Udviklingen af endotermi hos fugle og pattedyr er en vigtig overgang under hvirveldyrs evolution, hvilket giver et ekstraordinært eksempel på evolutionær konvergens mellem grupper, afgørende for deres udbredte geografiske udbredelse og økologiske succes. Selvom flere grupper af hvirvelløse dyr og hvirveldyr kan hæve deres temperaturer over omgivelserne, evnen til at opretholde høj og konstant kropstemperatur via endogen varmeproduktion i hvile er eksklusiv for fugle og pattedyr alene. Observationen forklarede deres større mobilitet, udholdenhed og tolerance over en række forhold. Imidlertid, denne strategi er energisk kostbar og har ingen virtuelle spor i fossiloptegnelsen, derfor forbliver tempoet og metoden for endotermi kontroversiel under hvirveldyrs evolution.
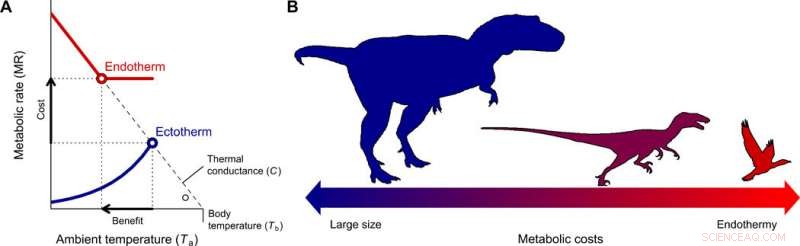
Udviklingen af endotermi og miniaturisering i theropod-slægten, der fører til fugle. (A) Fordelene ved at skifte fra ektotermi til endotermi for forskellige kropsstørrelser blev kvantificeret med Scholander-Irving-modellen, som beskriver hvordan en stigning i stofskiftet i hvile (omkostninger) øger den termiske niche Tb − Ta (fordel). Fordi der ikke er nogen termisk gradient mellem organismen og miljøet i fravær af varmeproduktion, denne kurve skærer abscissen ved Tb =Ta, når MR =0 (8). De solide blå og røde linjer viser de metaboliske kurver for en typisk ektoterm og endoterm, henholdsvis, og de åbne symboler viser den maksimale termiske gradient Tb − Ta mulig med hvilestofskiftehastigheder, brugt i vores model (lign. 2). (B) En reduktion i kropsstørrelse, i overensstemmelse med den, der er beskrevet fra forfædres theropoder til basalfugle (22), udgør den evolutionære vej til mindste modstand, da energiomkostningerne ved at være stor ombyttes med dem ved at være endoterme. Kredit:Science Advances, doi:10.1126/sciadv.aaw4486
For at forstå oprindelsen og årsagen til endotermi under udviklingen af fugle og pattedyr, forskerne overvejede to grundlæggende spørgsmål. Hvad er omkostningerne og fordelene ved denne strategi sammenlignet med ektotermi? Og hvilke forhold begunstigede en overgang til endotermi? Rezende et al. besvarede spørgsmålene ved hjælp af Scholander-Irving-modellen for varmeoverførsel, brugt til at studere termoregulering i endotermer i mere end 60 år. Forholdet blev sjældent brugt til ektotermer, men da alle levende organismer producerede endogen varme, modellen forblev anvendelig under termisk stabil tilstand. De gjorde denne afgørende antagelse for at undgå at bruge komplekse dynamiske modeller, der ofte anvendes til ektotermi, hvilket kunne gøre de foreliggende analyser vanskelige.
De kvantificerede omkostningerne ved endotermi som masseuafhængige energiudgifter, hvor fordelene omfattede større mobilitet og fourageringseffektivitet, undgåelse af rovdyr, tolerance over for og kolonisering af en lang række miljøforhold, med øgede vækstrater og homeostase. Rezende et al. kvantificerede den termiske niche, som organismer kunne besætte, og dens ekspansion for at estimere nettofordelen ved endotermi. De beregnede derefter cost-benefit ved at vedtage en endoterm livsstil med reference til den ektotermiske forfader og endoterme efterkommer. Som oprindeligt foreslået af den evolutionære biolog Bran K. McNab, da forskerne gentog disse beregninger med nøjagtige skøn over kropsstørrelsen, resultaterne viste, at mindre størrelser reducerede energiomkostningerne for at udvikle sig mod endotermi.
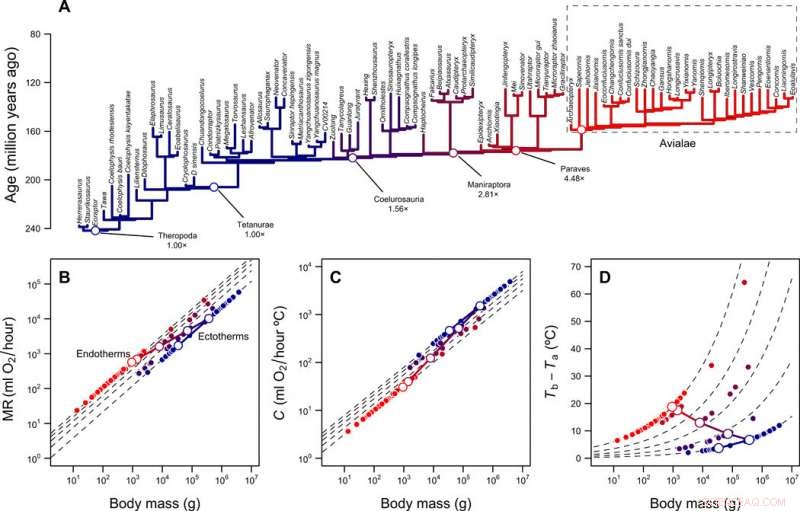
Rekonstruktion af metaboliske niveauer og termisk niche af theropoder. (A) Theropod fylogeni med grene farvekodet i henhold til rekonstruerede metaboliske niveauer. (B) Skalering af stofskifte versus kropsmasse for ektotermer (MR =0,68 masse 0,75) og endotermer (MR =3,4 masse 0,75) og den forudsagte bane for fuglestammens linje under overgangen fra ektotermi til endotermi. Stiplede linjer viser foldforskelle mellem ektotermer og endotermer (1× til 5×); åbne og lukkede symboler viser rekonstruerede værdier for fuglestammens slægt og spidserne af fylogenien, henholdsvis. (C) Skalering af termisk ledningsevne C og kropsmasse for ektotermer (C =2,5 masse 0,5) og endotermer (C =1,0 masse 0,5), fold forskelle fra 2,5× til 1×. (D) Termisk gradient og foldforskelle beregnet med lign. 1 (i publikationen) og værdierne i (B) og (C). Log-log lineære baner, der forbinder MR og C af den ektotermiske forfader og den endoterme efterkommer, såvel som den resulterende bane i termisk gradient, er vist med kontinuerlige linjer. Kredit:Science Advances, doi:10.1126/sciadv.aaw4486
Holdet undersøgte, hvordan denne varmeoverførselsmodel kombineret med fylogenier og kropsstørrelsesrekonstruktioner, kaste lys over udviklingen af endotermi hos fugle og deres theropod-forfædre. Rezende et al. estimeret omkostningerne ved at udvikle endotermi langs fuglestammen ved hjælp af rekonstruerede forfædres kropsstørrelser, baseret på fossiler. For at kvantificere energiomkostningerne inden for alternative scenarier simulerede de udviklingen af kropsstørrelse langs slægten og opnåede fordelingen af omkostninger pr. grad i modellen. De antog en urettet Ornstein-Uhlenbeck (OU) model mellem en masse på 10 g og 100, 000 kg med en gennemsnitlig udviklingshastighed svarende til dem, der er rapporteret for theropoder. Simuleringerne indikerede et markant fald i energiomkostningerne pr. grad med miniaturisering. Forskerne forklarede de reducerede omkostninger ved at bruge to fænomener.
Først, udvidelsen i termisk niche baseret på en stigning i metabolisk hastighed (MR) var uforholdsmæssigt højere i større ektotermer på grund af deres evne til at opretholde en høj kropstemperatur (T b ). Dette blev ledsaget af en relativt lav masseuafhængig metabolisk hastighed, på grund af initial homeothermi (stabil termoregulering). Derfor, de bemærkede, at større startstørrelsen af den ektotermiske forfader, billigere overgangen til endotermi. Sekund, under miniaturisering, dyrene byttede energiomkostningerne ved at være store for at være endoterme. Resultaterne forklarede, hvordan de høje energiomsætningshastigheder udviklede sig, uanset deres indvirkning på fødevare- og vandbehov. På trods af iboende variation i ressourcetilgængelighed i evolutionær tid - mindre størrelser og højere energiomsætningshastigheder blev systematisk favoriseret i denne afstamning. Resultaterne af arbejdet stemte kvalitativt overens med andre modeller om udviklingen af endotermi.
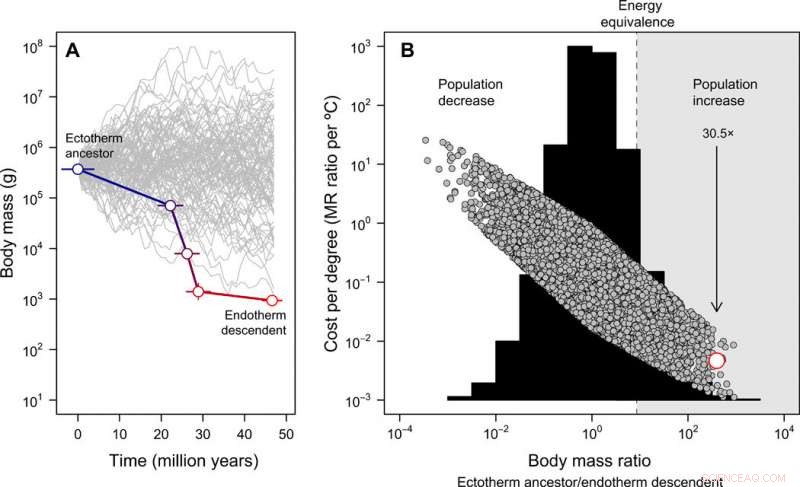
Udvikling af kropsstørrelse og cost-benefit ved endotermi. (A) Miniaturiseringen fra Tetanurae til basale fugle udledt fra fossiloptegnelsen, i modsætning til 100 simulerede størrelsesbaner startende fra den samme forfædres kropsstørrelse til illustrative formål (bemærk, at for den efterfølgende fulde nulmodel, den forfædres kropsstørrelse tillades at variere). Fejlen repræsenterer SD i rekonstruerede værdier på tværs af 20 kandidattræer. (B) Frekvensfordelingen af kropsmasseforhold opnået over 10, 000 simulerede kropsstørrelsesbaner (histogram) og energiomkostningerne til at udvikle endotermi udtrykt pr. grad Celsius (lign. 2 i publikationen) under denne nulmodel (grå symboler). I dette tilfælde, den forfædres kropsstørrelse blev opnået fra en ensartet fordeling på mellem 10 g og 100, 000 kg. Det empiriske skøn i fuglestænglen er vist med rødt. Den region, hvor en reduktion i kropsstørrelsen ville kompensere for energiomkostningerne ved at udvikle endotermi, gør det muligt for befolkningen at stige i et scenarie med konstante ressourcer, er fremhævet med gråt. Pilen viser den forventede befolkningstilvækst, givet den observerede kropsstørrelsesreduktion i fuglestænglen efterhånden som endotermi udviklede sig. Disse analyser indikerer, at energiomkostningerne til at udvikle endotermi reduceres med miniaturisering og, som resultat, befolkningsstørrelsen kan være steget på trods af de metaboliske omkostninger ved en endoterm livsstil. Kredit:Science Advances, doi:10.1126/sciadv.aaw4486
Størrelsesreduktionen i fuglestammens afstamning, nøje matchet den teoretiske vej for mindste modstand for at endotermi kan udvikle sig, derefter Rezende et al. rekonstrueret, hvordan fænomenet kunne have udspillet sig i therapod-fylogenien. Processen indikerede stigningen i MR (metabolisk hastighed), der spænder over det meste af Tidlig-Middel Jura (ca. 180 til 170 millioner år siden), involverer terapeutgrupper, hvor forekomsten af protofjer og fjer allerede var allestedsnærværende. De foreslog også meget forskellige metaboliske hastigheder på tværs af de samtidige afstamninger af Coelurosauria, Maniraptora og Paraves, som kunne redegøre for fremkomsten og diversificeringen af disse grupper i den sene jura periode. Arbejdet kunne også forklare den unormalt høje mangfoldighed af Coelurosauria ved mellemliggende kropsstørrelser (30 til 300 kg) sammenlignet med andre dinosaurgrupper. Forskerholdet demonstrerede yderligere en nichefyldende model af adaptiv stråling i mesozoiske dinosaurer med exceptionelle hastigheder af reduktion af kropsstørrelse i fuglestammens slægt (dvs. inden for basalknuder i Coelurosauria og Paraves), imidlertid, de foreslog ikke en passende evolutionær hypotese til at redegøre for resultatet.
På denne måde Enrico L. Rezende og kolleger observerede to exceptionelle fænomener under fuglenes udvikling; (1) en vedvarende (men ikke nødvendigvis gradvis) miniaturisering, der spænder over millioner af år og (2) fremkomsten af endotermi. De brugte rekonstruktioner til at foreslå samtidig udvikling af endotermi med et fald i størrelse langs fuglestammen, som også tidligere foreslået for pattedyr. De tilsvarende terapeutiske klader udviste et helt spektrum af MR. Fortolkningerne indikerer, at endotermi gik forud for udviklingen af flyvning, og den markante reduktion i kropsstørrelse under udviklingen af fuglestængel-slægt blev ledsaget af et stort skift i metaboliske niveauer.
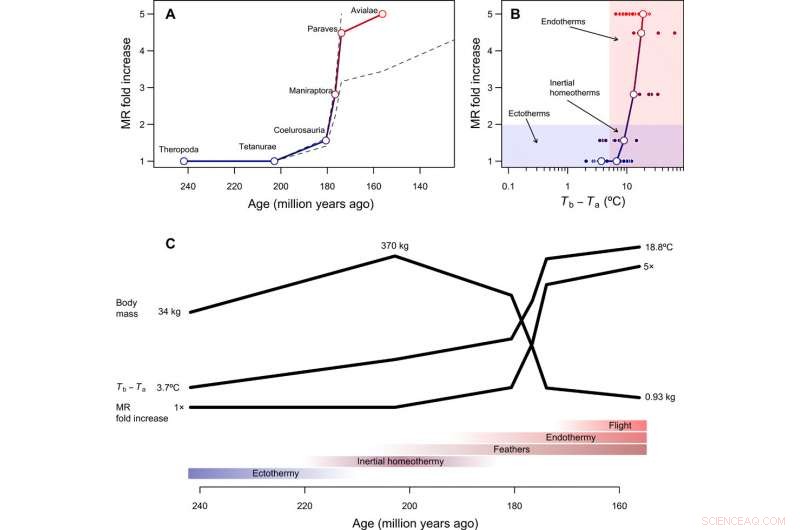
Tempo og tilstand i udviklingen af endotermi. (A) Rekonstrueret tidsmæssigt forløb af metabolisk udvikling i fuglestammen, med stiplede linjer, der viser, hvordan rekonstruktioner ændrer sig under antagelse af, at enten Paraves eller Neornithes var fuldt endoterme i stedet for basalfuglen [til beregninger med Neornithes, vi antog en kropsstørrelse på 150 g baseret på estimater for Vegavis og et tidsestimat på 100 Ma siden]. Den foldede stigning i MR blev beregnet ved at dividere den rekonstruerede MR under overgangen til endotermi med den forventede MR for en ektoterm af tilsvarende størrelse og er derfor dimensionsløs og uafhængig af kropsstørrelse. (B) Den evolutionære vej for mindste modstand fra ektotermi til endotermi inkluderer inerti homøotermi som en overgangsfase, efterfulgt af en stigning i stofskiftet samtidig med en reduktion i størrelse. (C) Hypotetisk sekvens af evolutionære overgange i fuglestammen, som kombinerer resultater fra denne undersøgelse med fylogenetiske rekonstruktioner af epidermale strukturer og kapacitet til aktiv flyvning (se hovedteksten). Kredit:Science Advances, doi:10.1126/sciadv.aaw4486
Det foreslåede scenarie forklarede, hvorfor theropoder systematisk favoriserede en dyr livsstil (højere energiomsætningshastighed) på trods af dets energiomkostninger, og vedvarende miniaturisering forud for fuglenes oprindelse. Desuden, en tilstand af inerti homøotermi udgjorde en nødvendig overgangstilstand med lave metaboliske omkostninger. Hvis store forfædres theropoder var fysiologisk forpligtet til homøotermi, Rezende et al. forvent derefter, at de er gået fra ektotermisk metabolisk allometri (forholdet mellem kropsstørrelse og form, anatomi, fysiologi og adfærd) med miniaturisering. I evolutionær tid, det er rimeligt at forvente, at slægter udnytter nyåbnede nicher og til sidst diversificerer. Resultaterne af dette arbejde er foreløbige; de fremhæver blot, at der sandsynligvis er mere til udviklingen af endotermi hos arkosaurer, dinosaurer og nutidens fugle. Resultaterne danner en arbejdshypotese, der i fremtidige undersøgelser kan kombineres med tidligere analyser af udviklingen af størrelse og andre karakteristika i fuglestammeafstamning for at danne veldefinerede fortolkninger af tidsmæssige sekvenser under centrale evolutionære overgange.
© 2020 Science X Network
 Varme artikler
Varme artikler
-
 At løse et gammelt mejerimysterium kan hjælpe med at helbrede moderne madsygdommeAt studere kulturen af mongolske hyrder kan hjælpe med at knække mysteriet om, hvorfor mennesker begyndte at indtage animalsk mælk, før befolkningerne udviklede sig til at være i stand til at fordøj
At løse et gammelt mejerimysterium kan hjælpe med at helbrede moderne madsygdommeAt studere kulturen af mongolske hyrder kan hjælpe med at knække mysteriet om, hvorfor mennesker begyndte at indtage animalsk mælk, før befolkningerne udviklede sig til at være i stand til at fordøj -
 Forskere borer ned til kernen af velvære på verdensplanKredit:Pixabay/CC0 Public Domain Forskere ved SAHMRI og Flinders University har udført den største meta-analyse nogensinde af velværestudier fra hele verden for at besvare spørgsmålet, Hvad er den
Forskere borer ned til kernen af velvære på verdensplanKredit:Pixabay/CC0 Public Domain Forskere ved SAHMRI og Flinders University har udført den største meta-analyse nogensinde af velværestudier fra hele verden for at besvare spørgsmålet, Hvad er den -
 Kønsnormer påvirker holdninger til homoseksuelle mænd og lesbiske kvinder globaltKredit:CC0 Public Domain Homoseksuelle mænd og lesbiske kvinder har ofte været mål for fordomme og endda vold i samfundet. For bedre at forstå, hvad der former disse holdninger og fordomme, Maria
Kønsnormer påvirker holdninger til homoseksuelle mænd og lesbiske kvinder globaltKredit:CC0 Public Domain Homoseksuelle mænd og lesbiske kvinder har ofte været mål for fordomme og endda vold i samfundet. For bedre at forstå, hvad der former disse holdninger og fordomme, Maria -
 Hvad er engelhårfænomenet?Ingen videnskabsmand har været i stand til at samle englehår, men ud fra beskrivelserne, de gætter på, at det kan være spindelvæv. Photos.com/Thinkstock Her er et tip:Når du skriver om et fænomen, de
Hvad er engelhårfænomenet?Ingen videnskabsmand har været i stand til at samle englehår, men ud fra beskrivelserne, de gætter på, at det kan være spindelvæv. Photos.com/Thinkstock Her er et tip:Når du skriver om et fænomen, de
- Hvordan vi opdagede 840 mindre planeter hinsides Neptun - og hvad de kan fortælle os
- Undersøgelse for at undersøge sammenhængen mellem græsarealer og jordens biodiversitet
- Første radiodetektering af en ensom planetskive viser ligheder mellem stjerner og planetlignende ob…
- Forskere håber, at nogle genstande overlevede Brasiliens museumsbrand
- NASAs GRACE:Hvad forskere har lært af vand i bevægelse
- Silicon giver midler til at styre kvantebits til hurtigere algoritmer